
Amongst the most challenging biological enigmas is how avian flight evolved. This can be attributed to the contradicting features of the oldest accepted avian, the Archaeopteryx. Extant birds also possess anextensive diversity with varied flying mechanisms, which further complicates the conundrum. Historically, there has been a strong debate around two hypotheses, namely the Cursorial and the Arboreal models. However, lack of concrete evidence for both suggests an evolutionary route which integrates both models. Wing-inclined Assisted Flight hypothesis and the Pouncing Proavis model are both possible modifications of the former two explanations. Nonetheless, the advent of flight, like most evolutionary events, have resulted in the creation of a plethora of species.
The Cursorial Model
The cursorial model implies that the evolution of flight occured through the utilization of short jumps by running bipedal terrestrial theropod. Albeit temporarily, the upliftment from the ground provided theropods with an apparently effective predation mechanism. Through motions similar to wing strokes in extant birds, the theropods employed extended and flattened forelimbs to enhance body oriental control and progressively increase lift. Flight is achieved when the residual lift is greater than the individual’s bodyweight.
Alongside the flapping of forelimbs, the hindlimbs play a significant supporting role, particularly in garnering sufficient running velocity and thrust (Norberg, 1985). Supporting evidence for this hypothesis lies in the long and erect hindlimbs of the Archaeopteryx, emphasizing their terrestrial nature. However, it must be noted that this evidence is severely depleted.
The Model Lacks Evidence
A prominent argument for this model is the hypothetically calculated 4 metres per second velocity gap between the minimum jumping and maximum running speeds of the Archaeopteryx. The velocity gap implies an inability to generate sufficient force for flight. However, these calculations are based on the assumption that the hindlimbs were solely associated with running speeds. The effects of forelimb flapping were not considered in these models. A reworked model, which integrates these factors, estimates that the maximum running speed of Archaeopteryx in fact be 1.8m/s greater than required minimum jumping speed. This model, therefore nullifies this criticism of the cursorial model. (Burgers and Chiappe, 1999)
Secondly, the debate regarding the evolution of symmetrical forelimb movement in territorial precursors remain unresolved. A running theropod requires the simultaneous and symmetric movement of forelimbs to generate sufficient lift. However, such forelimb movement also elevates the drag encountered by the theropod. The associated reduction in speed would have had a severe reduction in thrust. (Nudds and Dyke, 2009) Due to this limitation, it is presumed that a competitive pressure would be required for the evolution of such forelimb motion.
The Wing Inclined Assistance Running Hypothesis
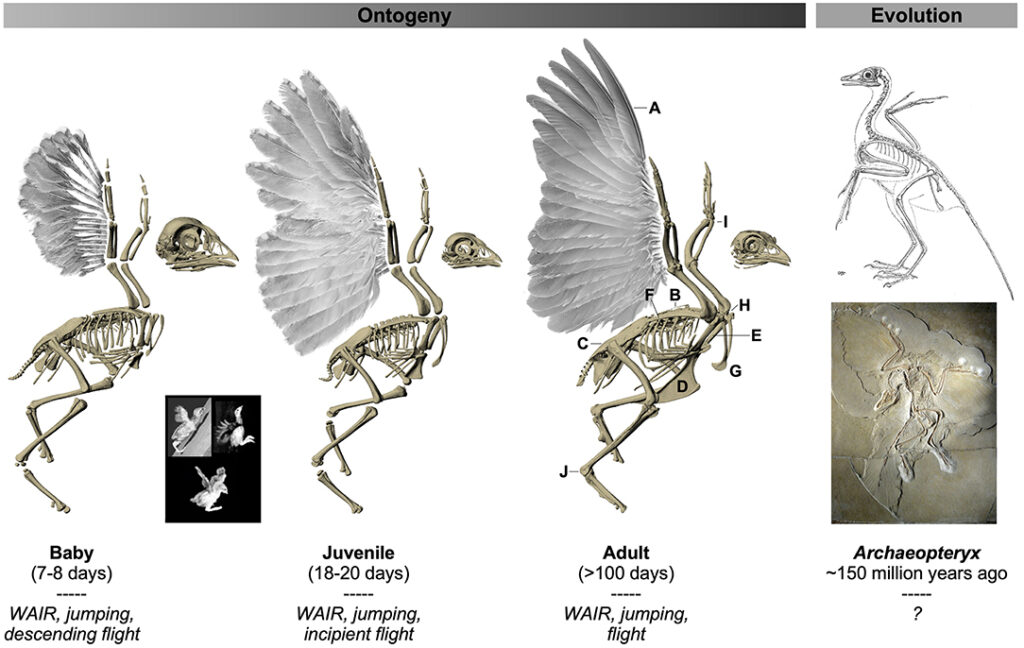
The Wing Inclined Assistance Running Hypothesise (WIAR) is a potential justification of such development. This theory suggests that avian precursors were required to run up inclined paths, such as tree trunk, to avoid predation. Aerodynamic adaptations, such as forelimb modifications, could enhance acceleration ability. However, the lack of shoulder mechanisms in early birds, suggests the inability to conduct the required upstroke movement. (Senter, 2006). Nonetheless, a more comprehensible justification of symmetric flapping is provided by the Arboreal Flight Hypothesis.
This theory suggests that the Archaeopteryx was partially arboreal in nature and glided between trees. Wings were essential for balancing purposes. It is assumed that the evolution of larger feathers leads to a paradigm shift towards powered flight.
The Arboreal Hypothesis
The arboreal theory provides for a more energy efficient model than the cursorial hypothesis. The costs of tree climbing were negated by the efficiency of gliding. Considering a higher starting point, greater speeds are generated during flight. This allows for gliding aerodynamic to act upon the organism, hence reducing energy dissipation. Additionally, the theory simplifies, although not resolve, the practicality of symmetric forelimb movement. The inefficiencies of simultaneous running and flapping would be negated by arboreal based flight. (Dudley and Yanoviak, 2011)

But limited evidence……….
The arboreal hypothesis is partially supported by morphological evidence. One such example is the Archaeopteryx hallux, which is the modified first digit of a foot.Contrary to earlier perceptions, the hallux was not pointed posterior to remaining digits, allowing for perching behaviour. This adaptation compensates for the inflexible ankle of birds, which would otherwise prevent climbing behaviour. However, the interpretation of the Archaeopteryx hallux fossils could be influenced by disarticulation. The earliest definitive evidence of a reversed hallux is in the fossils of substantially younger avian Sapeornis. Another feature in the Archaeopteryx fossils which bolsters arboreal ability is the manus, an important forelimb segment used for gripping. (Mayr, Pohl, Hartman and Peters, 2007).
Meanwhile, evidence of the gliding origin of flight has been correlated with appearance of elongated tibia feathers in early avian specimen. This evidence supposedly invalidates the cursorial theory. This is because the feathers could potentially impend terrestrial locomotion, which would lead its disfiguration. However, the feathers would also be exposed to similar magnitudes of defacement in an arboreal environment due to extensive manoeuvring within branches. (Dececchi and Larsson, 2011). The ambiguity of such morphological features of avian percussors prevent the validation of the Arboreal Hypothesis of Flight Evolution.
The Pouncing Proavis Model
On the basis of the obscurity of both models, a third hypothesis, the Pouncing Proavis Model, has been proposed. The postulation encompasses both of the former theories in attempt to justify morphological adaptation. The basis of model lies in the predatory nature of avian precursor. These predators, through the use of raptorial hindlimbs, employed hoisted platforms for ambuscades.
The model suggests that specimen originally possessed drag based mechanisms which evolved into more sophisticated system involving aerodynamic lift. This led to enhanced control and increasing predation range. It also implies a probably concurrence of primitive pounces ( Microraptor) and advanced fliers (Confuciusornis) in different flying niches. It must be noted that the theory is still in its rudimentary stages. (Garner, Taylor and Thomas, 1999)
In conclusion, the evolution of avian flight remains ambiguous. Both of the more renowned theories, despite their merits, remain inconclusive. There is a growing inclination towards the possibility of the integration of both hypotheses. The Pouncing Proavis Model, though not explore sufficiently, is certainly in lieu with this more practical approach. Regardless, the advent of flight has allowed birds to adapt to changing environments.
Help us Help Them! Think Wildlife Foundation is a non profit organization with various conservation initiatives. Our most prominent campaign is our Caring for Pari intiative. Pari is a rehabilitated elephant at the Wildlife SoS Hospital. 25% of the profits from our store are donated to the elephant hospital for Pari. Other than buying our wonderful merchandise, you could donate directly to our Caring For Pari fundraiser.